
|
|
はじめに 飽食の時代といわれてから久しく、街には食料品があふれています。私が小学生の頃の給食は、今では想像もできないまずいパンと脱脂粉乳だったことを思い浮かべると、隔世の感があります。しかし、カロリーで計算した日本の食糧自給率が、実は四〇%足らずであるということをご存じでしたでしょうか?いわゆる先進国と呼ばれている国々と比較すると、極端に低い値です。最近の、地球規模での環境変化や、輸入農産物・食品の安全面を考えると、大変心許ない状況にあります。生活する上で最も重要な食料の生産を、真剣に考え直す時期にきていると考えています。一方、地球全体で収穫できる穀物では、約五〇億人を養う程度と言われています。この数字は一九八七年の人口と同じ位で、すでに食料は不足しています。植物は、十七種類の無機元素を駆使して、全ての有機物を生合成します。人や動物の生存は、植物が作った有機物に依存しているといっても過言ではありません。この中で、植物が多量に必要で、最も不足するのが窒素です。 イネの研究の重要性とは イネは三大穀物の一つであり、アジア地域で全体の九〇%以上が生産されています。他のコムギやトウモロコシに比較して、お米は粉にしなくても食べられる穀物です。研究材料として見た場合、ゲノム(遺伝子を含む全DNAのこと)のサイズがトウモロコシの六分の一、コムギの四〇分の一と小さく、二〇〇四年十二月にゲノムの塩基配列は完全解読されていて、農学や生物学においてモデル生物の一つとして重要な位置にあります。研究に必要な分子資材や突然変異体の整備も、順次進められております。大きな利点として、イネの染色体構造はコムギやトウモロコシなど他のイネ科作物の構造と似ていることから、イネで得られる結果が他の作物に応用可能であり、波及効果が大きい点が上げられます。このような利点を生かし、私どもの研究室では、イネを用いて、生産性や品質に最も影響をもつ窒素の同化反応や体内での利用機構の研究を二〇年以上継続して来ました。とりわけ、生産性向上や環境負荷を少なくする窒素利用効率の向上に関する分子機構の解明に取り組んできました。 窒素利用の分子機構について (1)根による無機態窒素の吸収と同化について 研究を開始した当時、分子生理学の研究は、動物に比較してはるかに遅れていて、代謝経路はある程度わかっていたものの、その素反応がどの細胞でなされているのかさえ、わからない状況でした。特異抗体を用いた組織染色や免疫電子顕微鏡、また緑蛍光タンパク質などを用いてmRNAの発現場所と時期を特定し、器官や組織を構成する細胞は、代謝機能が異なることを見いだしました。 水田で成育するイネは、水田土壌が還元状態であることから最も還元が進んだ無機態窒素であるアンモニウムイオン(NH4+)を主な窒素源として用います。NH4+は、根のNH4+輸送に関わるタンパク質によって吸収され、主に根の表層の細胞群でグルタミンやグルタミン酸というアミノ酸に同化されたのち、導管を通って地上部に運ばれます。この際、同化反応に関わるのがグルタミン合成酵素(GS)とグルタミン酸合成酵素(GOGAT)です。イネには、GSが四分子種(GS1;1, GS1;2, GS1,3, GS2)、GOGATが三分子種(NADH-GOGAT1, NADH-GOGAT2, Fd-GOGAT)存在しています。一方で、NH4+は、施肥由来以外に、タンパク質や核酸の異化反応や二次代謝経路、光呼吸経路など、体内での複数の代謝反応によっても生成します。しかし、一般に体内のNH4+濃度は極めて低く、同化反応が迅速かつ効果的に進められていることを示します。根でのNH4+同化にはGS1;2とNADH-GOGAT1が重要な働きをしている結果が得られました。 (2)窒素利用は、収量に直接影響します。 さて、イネの穂や種子(お米)を形成するためにも窒素は重要な働きをしますが、穂に運ばれてくる窒素の約八〇%は、若い時期に葉(葉身と葉鞘)や茎で活躍していたタンパク質などが分解され、篩管を介して輸送されてきた窒素に由来することが、安定同位体の窒素で標識した研究から判明しました。田んぼをよく見ると、下の葉から黄色になっていくことがわかります。この際、篩管液の主な窒素形態はグルタミンとアスパラギンというアミノ酸であることがわかっています。アスパラギンはグルタミンから生合成されますので、老化器官では、まずグルタミンの合成が必要となります。また穂に到達したグルタミンは、グルタミン酸に変換された後に、多くのアミノ酸が合成され、貯蔵タンパク質を始めとする重要な物質の素材となります。遺伝子発現の予備的な解析から、老化器官ではGS1;1が、また穂ではNADH-GOGAT1が重要であることが示唆されました(図1)。 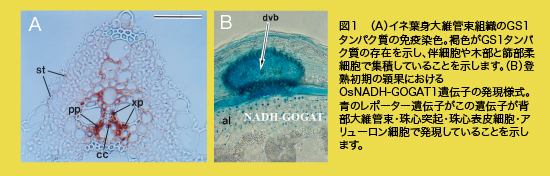
GS1;1の機能を証明するために、OsGS1;1遺伝子が破壊された変異体を用いて解析を行いました。この遺伝子破壊変異体は、成育が極めて遅く、登熟まで一ヶ月以上長くかかるとともに、背丈が半分程度、また小さな穂にほとんど中身が詰まっていない種子をつける表現型(見た目の性質)を示すことが判明しました(図2)。この表現型は、OsGS1;1遺伝子を変異体に再導入することで正常になることから、OsGS1;1遺伝子機能欠損によるものであることを結論づけることができました。 
窒素代謝と炭素代謝は、密接に連携をとっています。この変異体で、代謝物がどのように変化するのか、代謝物を一斉に調べた結果、窒素代謝と有機酸代謝に関連した物質が極端に低下し、炭素代謝関連物質が増加してバランスが大きく崩壊していることも判明しました。従って、GS1;1は、生産性を決定する窒素転流機構で重要な機能をしていることがわかりました。 窒素利用効率の向上を目指して 古くから、経験的に窒素の供給を抑えると、根の伸長が促進されることがわかっています(図3)。NH4+の供給濃度に対応して根の伸長を促進する原因遺伝子を、量的形質を決定する遺伝子座(QTL)を利用した解析から突き止めました。この遺伝子は、ある種の転写活性化因子をコードしており、根の伸長と同時にNH4+吸収量を促進する効果を持つことが判明しました。日本型イネでは、インド型イネよりもこの効果が小さいこともわかりました。今後、このような遺伝子を活用し、環境負荷が少なくてすむイネの開発を目指して、研究を進めていきたいと思います。 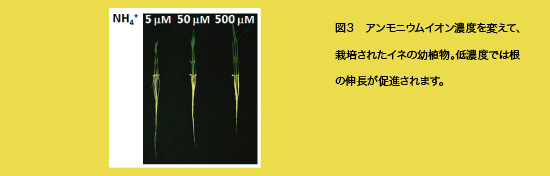
|
 |
山谷 知行(やまや ともゆき)
1950年生まれ 東北大学大学院農学研究科 教授 専門/植物細胞生物学 関連ホームページ/http://www.agri.tohoku.ac.jp/cellbio/index-j.html |


